El cuerpo humano es una máquina de vida, y a pesar de la necesidad de cuantificar y clasificar, todo individuo es más que el conjunto de lo que se puede medir, describir y comprender, incluso cuando esto se hace con la intención de ayudarlo.
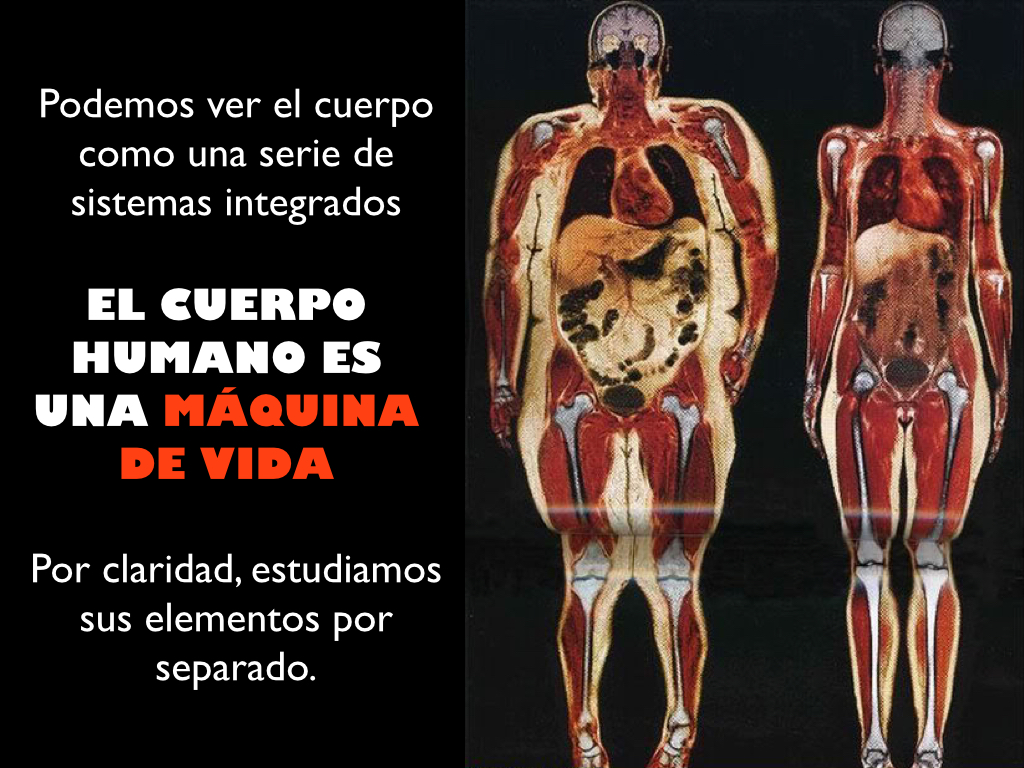
Podemos ver el cuerpo humano como una serie de sistemas integrados y por claridad, estudiar sus elementos por separado, pero sin olvidar que se trata de órganos individuales (asociación de diferentes tejidos) morfológicamente unidos, que crean sistemas morfológica y funcionalmente interdependientes donde los planos estructurales del organismo nos ayudan a entender mejor esta organización: Célula <-> Tejido <-> Órgano <-> Sistema <-> Organismo. Las células especializadas se agrupan formando tejidos para realizar funciones en nuestro organismo, siendo los cuatro tejidos que se distinguen en los mamíferos: nervioso, muscular, conjuntivo y epitelial.

En los tejidos, según los libros de histología, además de células, también hay un “material inerte”, la matriz extracelular (MEC), compuesta por sustancia fundamental, fibras y fluido extracelular, siendo la célula la encargada de producir y secretar al espacio extracelular los componentes de la misma (Gartner y Hiatt, 2011; Gartner y Hiatt, 2014; Thibodeau y Patton, 2007; Tortora y Derrickson, 2011). Según Gray’s Anatomy (Standring, 2008), este término se aplica a la suma total de sustancias extracelulares de los tejidos, un sistema de fibrillas proteicas conocidas por proteoglucanos capaces de captar agua.
Hoy en día se sabe de la importancia de la MEC, implicada no solo en el soporte, sino también en el movimiento, desarrollo, morfología y función de las células desde el principio de la vida, durante el desarrollo del embrión, donde la migración de las células está condicionada por moléculas de la MEC como la fibronectina, laminina, colágeno (tipo IV) los proteoglucanos y el sulfato de condroitina a las que se unen las células mediante las integrinas (proteinas de unión), permitiendo o inhibiendo su migración (Carlson, 2009) constituyendo así un medio iónico hidratado y poroso, que hace posible el que los metabolitos y nutrientes puedan difundirse con facilidad (Standring, 2008).

Hay que destacar que a pesar de encontrar textos que todavía presentan a la MEC como material inerte, la visión actual de este entramado molecular va cambiando, y como indica Myers (2015) citando a James Oschman en referencia a su libro Energy Medicine (Oschman, 2000) se empieza a considerar la MEC como la matriz viva:
«la matriz viva es una red ″supermolecular″ continua y dinámica que se extiende hasta el último rincón del cuerpo: una matriz nuclear dentro de una matriz celular incluida en una matriz de tejido conjuntivo. En esencia, cuando tocamos un cuerpo humano, estamos tocando un sistema íntimamente interconectado compuesto por la unión de la práctica totalidad de las moléculas del organismo».
Como afirma Mecham (2011), la comprensión de la función de la MEC ha evolucionado del concepto de “tejido conectivo” estático que mantiene unido y conecta todo, hacia el de un biomaterial dinámico que proporciona fuerza y elasticidad interactuando con los receptores de la superficie celular y controlando la disponibilidad de los factores de crecimiento. Esto plantea la necesidad de incorporar la biología celular para poder entender completamente la MEC. Esta idea ha sido reforzada por otros autores en publicaciones recientes (Alberts, Johnson, Lewis, Morgan, Raff, Roberts y Walter, 2015) donde se expone que la MEC es más que un andamio pasivo que proporciona soporte físico cumpliendo un rol activo y complejo, regulando el comportamiento de las células que la tocan, habitan en ella o se desplazan a través de su entramado, influenciando la supervivencia, desarrollo, migración, proliferación, forma y función.
A diferencia de otros tejidos, en el tejido conectivo hay una gran proporción de MEC (Thibodeau y Patton, 2007) que da soporte y cabida a las células del sistema (fibroblastos, adipocitos, células plasmáticas, macrófagos, linfocitos, mastocitos) mediante una estructura que proporcionan las fibras (colágeno, elastina, reticulina) que se entrelazan e interactúan las unas con las otras en un entorno que determina las cualidades del tejido (según sea líquido, semiliquido, gelatinoso o calcificado), compuesto por la sustancia fundamental o matriz amorfa y el fluido extracelular. (Gartner y Hiatt, 2011; Gartner y Hiatt, 2014).
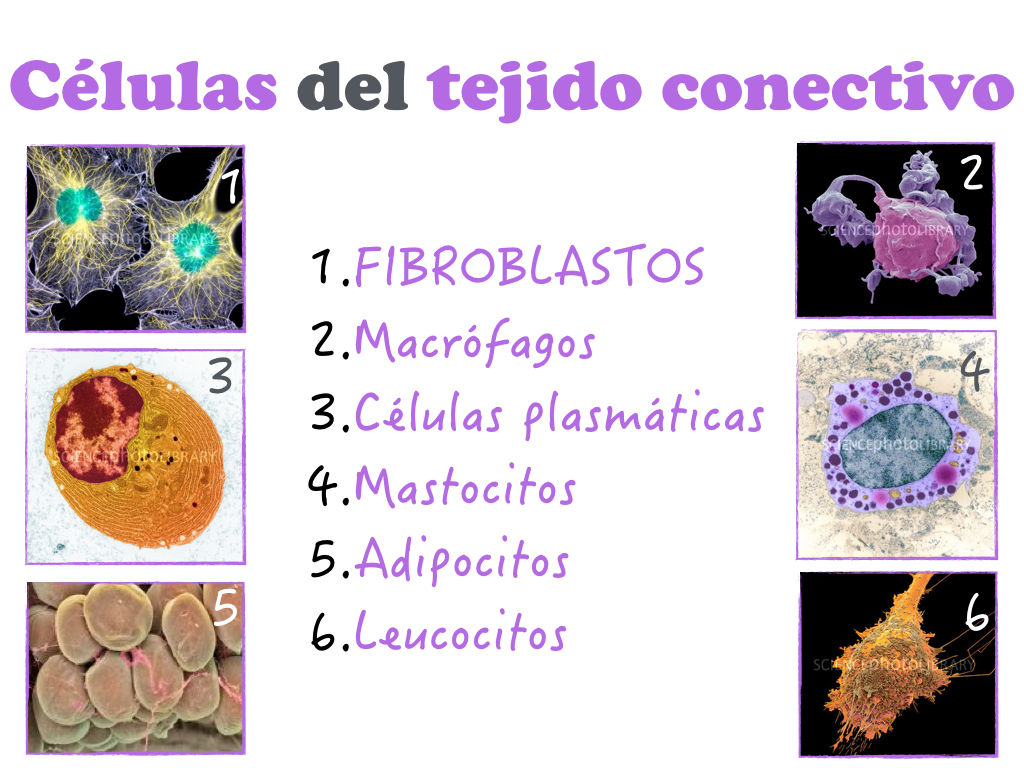

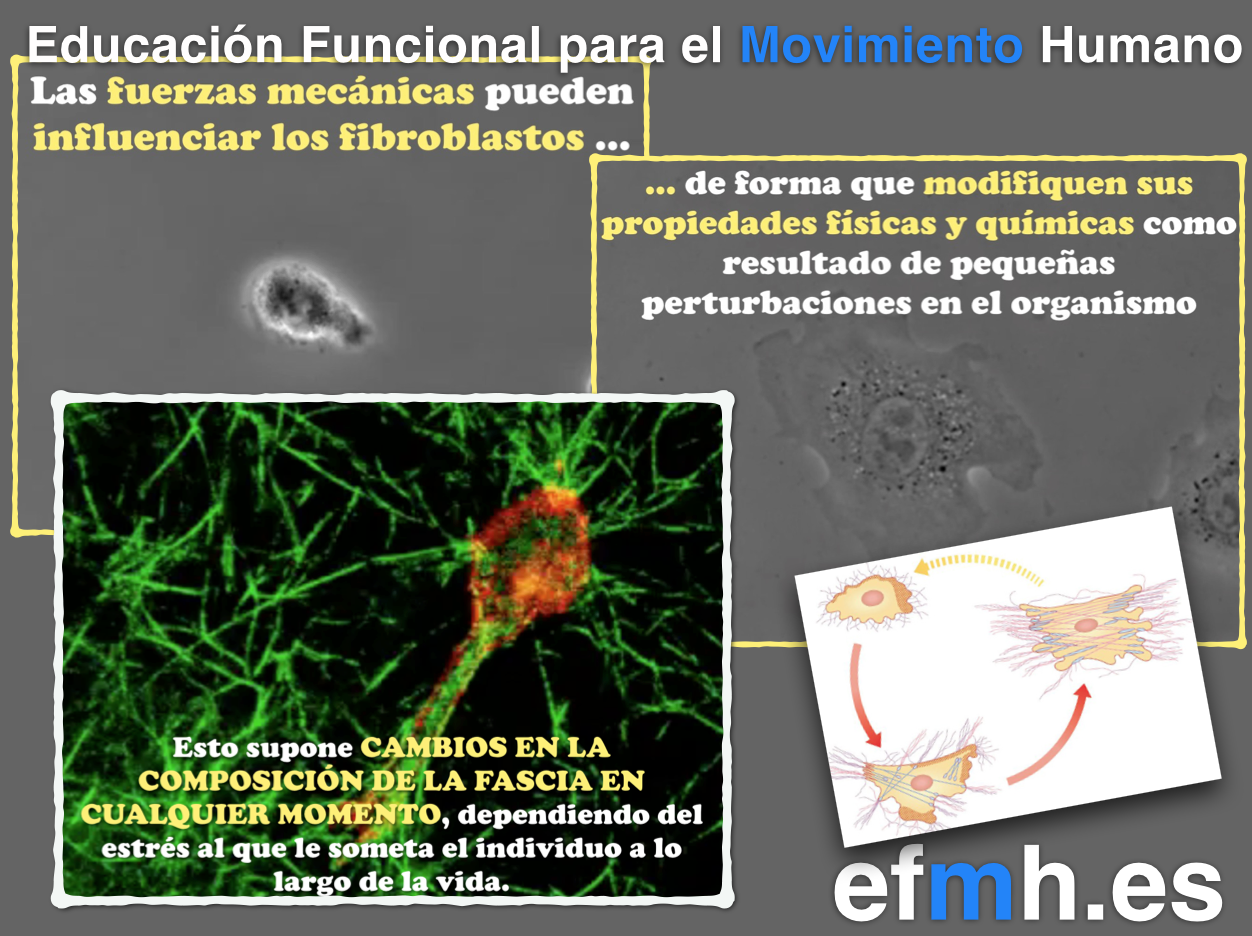
Como hemos visto, el tejido conectivo es algo más que un simple pegamento. Podemos decir de él que es el ingrediente que nos da la forma y a su vez, el componente que forma cada pieza de nuestro organismo, donde los mecanismos de unión y cohesión celular gobiernan la arquitectura, la forma, la fuerza y la organización de los diferentes tipos de células, unidas por interacciones directas o a la MEC. En este caos fibrilar debe primar la coherencia para poder soportar y responder las fuerzas tanto internas como externas (Alberts et al., 2015). Las células y por lo tanto tejidos que forman, se adaptan, cambian de forma con cada movimiento y se adaptan a cada situación tanto estática como dinámica del organismo.

Sus propiedades dinámicas dependen de su arquitectura biológica y aquí hablamos de una organización tridimensional que debe su fuerza, resistencia y adaptabilidad a la transmisión de fuerzas a través de la red molecular más importante de nuestro organismo que, a su vez, encuentran el equilibrio gracias al principio de la biotensegridad…, la red fascial
El significado funcional de la arquitectura del tejido conectivo y más concretamente de la red colagénica fascial se caracteriza por su capacidad para desestructurarse y permitir así la movilidad, así como su dimensión conectiva que permite interacciones mecánicas funcionales (entre los órganos o estructuras). En otras palabras, cuando hablamos de fascia, además de utilizar un término muy a la moda, hacemos referencia a un tejido que además de conectar, acompaña y hace posible el movimiento, siendo por lo tanto:
1) Capaz de adaptarse y cambiar su organización para permitir el movimiento (piensa en el deslizamiento de las fascias que envuelven los músculos, a nivel del gemelo por ejemplo, cuando caminas o al pasar de flexión plantar a dorsiflexión del pie. ¿Te habías parado a pensar cómo este deslizamiento es posible?).
2) Capaz de reorganizarse y unirse cuando se le necesita (esto es lo que permite la cicatrización de los tejidos dañados o la especialización de algunas zonas como los tendones que son en realidad una adaptación funcional de la fascia ante una demanda de tracción).
El concepto de tensegridad se basa en la coexistencia de fuerzas mecánicas, tensión continua y compresión discontinua, que interactúan para equilibrarse formando los binomios tensión – tracción y compresión – empuje (Heartney, 2009). Hablamos de fuerzas y por ende de energía, fuerzas/energía que podemos dibujar sobre un papel, sentir cuando nos movemos, percibir al observar un gesto, pero que aunque estén ahí no podemos ver…, fuerzas, energía, interacción, equilibrio.
Algo que caracteriza a las estructuras tenségricas es la capacidad de cambiar de forma y recuperarla gracias a sus propiedades viscoelásticas no lineares que hacen posible el auto equilibrado de las fuerzas, verdaderas estructuras dinámicas, formas siempre en tensión, siempre en equilibrio, en definitiva, estables (Juan y Tur, 2008; Skelton y de Oliveira, 2009; Scarr, 2014).
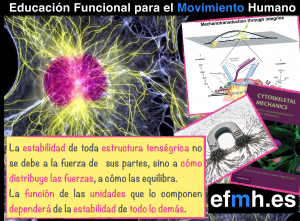
La estabilidad de toda estructura tenségrica no se debe a la fuerza de sus partes, sino a cómo gestiona el estrés mecánico al que es sometida, o dicho de otro modo, a cómo distribuye las fuerzas, a cómo las equilibra, a cómo encuentra el equilibrio estable (y dinámico de la materia viva) donde la función de las unidades que lo componen dependerá de la estabilidad de todo lo demás. (Juan y Tur, 2008; Scarr, 2014)
Si hablamos de formas tenségricas solo podemos hacerlo desde una visión de interacción tridimensional, introduciendo en nuestro vocabulario los conceptos de volumen y sinergia. Por ejemplo, cuando un globo que se infla, crece, crece y la distancia entre sus elementos aumenta a cada momento. Si esa distancia aumenta, las fuerzas “invisibles” que interactúan sobre ellos también lo hacen “me muevo luego existo”.
Extrapolando esta idea a nuestra naturaleza humana móvil y en constante cambio, tenemos un buen ejemplo que nos ayudará a entender la perspectiva global de toda postura o gesto que realizamos en nuestro día a día y las interacciones entre las fuerzas “invisibles”.
Una perspectiva global del movimiento nos enseña que en cada gesto existen elementos que se expanden acompañados de elementos en contracción. Tensiones que cambian y se reparten buscando el equilibrio del sistema, tantas veces alejado del equilibrio ideal, basándose en conceptos no lineares que entienden las sinergías entre los elementos que permitirá la transformación y el propio movimiento.
Si aplicamos el concepto de biotensegridad a la dinámica corporal, a su realidad de continuidad total, no podemos apartar la mirada ante la evidencia de la organización fascial a todos los niveles: superficial, profundo e interno (nivel visceral). Obviarlo supone no querer aceptar la realidad tridimensional de nuestra estructura, la realidad tridimensional del movimiento, afincarnos en la comodidad de teorías aprendidas de origen renacentista, desarrolladas cuando las mentes dibujaban en 2D.
El Profesor de la Universidad de Harvard, Donald Ingberg en sus publicaciones (Ingber, 2010; Sultan, Stamenović e Ingber, 2004) nos muestra como a nivel microscópico, la organización de la matriz extracelular (MEC) hace que la transmisión de fuerzas y tensiones llegue hasta el citoesqueleto de las células. Esto nos hace tener en cuenta el que toda fuerza mecánica que tiene lugar en nuestro organismo, se traduce en un proceso bioquímico, es decir que: cada gesto, cada movimiento, cada estímulo, cada sentimiento, cada acción mecánica o la simple y terrible “inacción” llegará hasta lo más profundo de nuestro ser, a cada unidad funcional de nuestro organismo, la Célula, desencadenando un cambio mediante el proceso de mecanotransducción.

Junto con la fascia, el tejido conectivo hace que los huesos conecten y funcionen correctamente, hasta el punto de que algunos autores definan la cápsula articular como una fascia especializada, creada a partir de la demanda mecánica multidireccional a la que se ven sometidos los tejidos de la zona y que consigue mediante este formato, aportar conexión y permitir la movilidad.
Reflexionando a partir de las ideas expuestas en este post sobre la MEC y su importante papel en la dinámica en constante cambio, adaptación y remodelación de los tejidos, es importante que cuando llevemos a cabo un trabajo de liberación miofascial, independientemente de si es antes o después de entrenar o pasear, independientemente de si buscamos una mejora del ROM, preparar/activar una zona o evitar la fatiga, tenemos que recordar que el entramado fascial no es tan solo miofascial, sino un órgano “tensotivomotor” (tensional – sensitivo – motor) implicado en los procesos microscópicos y macroscopicos del movimiento y de la vida.
Bibliografía:
Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., Walter, P (2015) Molecular biology of the cell. 6th ed. Ed. Garland Science.
Carlson, B.M. (2009) Embriología humana y Biología del desarrollo 4ºed Ed Elsevier.
Gartner L.P., Hiatt, J.L. (2011) Histología Básica. 1ª ed. Ed Elsevier.
Gartner L.P., Hiatt, J.L. (2014) Color Atlas and Text of Histology. 6ª ed. Wolters Kluwer Health.
Heartney, E. (2009) Kenneth Snelson: forces made visible. Massachusetts, Hard Press Editions.
Ingber, D. E (2010) From Cellular Mechanotransduction to Biologically Inspired Engineering. Annals of Biomedical Engineering, 38 (3), p. 1148-1161.
Juan, S.H., Tur, J.M.M. (2008) Tensegrity frameworks: static analysis review. Mechanism and Machine Theory 43, pp.859-81.
Mecham, R.P. (2011) The Extracellular Matrix: an Overview. Ed. Springer
Myers, T. W. (2015) Vias Anatómicas: Meridinos miofasciales para terapeutas manuales y del movimiento. 3ª ed Ed. Elsevier.
Oschman, J. (2000) Energy medicine. Edinburgh: Churchill Livingstone.
Scarr, G. (2014) Biotensegrity. The Structural Basis of Life. Handspring Publishing.
Skelton, R.E. y de Oliveira, M.C., Tensegrity Systems. Ed. Springer.
Standring, S. (2008) Gray’s anatomy. 40th ed Edinburgh: Churchill Livingstone.
Sultan, C., Stamenović, D., Ingber, D. E., (2004). A Computational Tensegrity Model Predicts Dynamic Rheological Behaviors in Living Cells. Annals of Biomedical Engineering, 32 (4), p. 520-530.
Thibodeau, G. A., Patton, K. T. (2007). Anatomía y fisiología. Ed. Elsevier.
Tortora, G. J., Derrickson, B. H., 2011. Principles Of Anatomy And Physiology (Tortora, Principles of Anatomy and Physiology). 13th ed. ed. Hoboken,NJ: Wiley.

